DFG founded research groups » Research Group Schmidt » Research
Research interests and techniques
We are interested in the mechanisms and the dynamics of information transfer between cortical neurons in the mammalian brain. In these processes the second messenger ion Ca2+ is of fundamental importance. Ca2+ triggers neurotransmitter release and regulates synaptic efficacy via ontogenetic and activity-dependent mechanisms of synaptic plasticity. We investigate synaptic transmission and plasticity on the cellular and molecular level, using techniques like paired patch-clamp recordings, two-photon (2P) microscopy and numerical computer simulations.
Synaptic transmission and plasticity
To lurk into neuronal communation we perform paired patch-clamp recordnigs in actue brain slices (Fig. 1). Qunatal synaptic release parameters are quantified by fluctuation analysis (Fig. 2) and synaptic Ca2+ signalling by dual-dye 2P- Ca2+ imaging (Fig. 3). Numerical computer simulations constrained by the experimental data are used to derive estimates about active zone topography (Fig. 4).
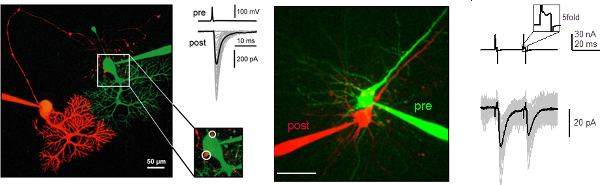
Figure 1. Paired recordings from connected Purkinje neurons in the cerebellar cortex (left; from Bornschein et al., J. Physiol., 2013) and from pyramidal neurons in the neocortex (right; from Bornschein et al., 2019).
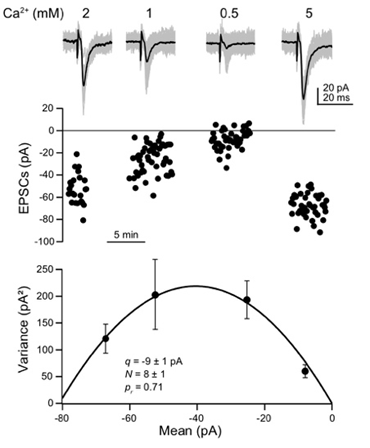
Figure 2. Fluctuations of EPSC amplitudes recorded from pairs of connected neurons at different extracellular Ca2+ concentrations can be used to estimate quantal synaptic release parameters (quantal size, release probability, and number of release sites).
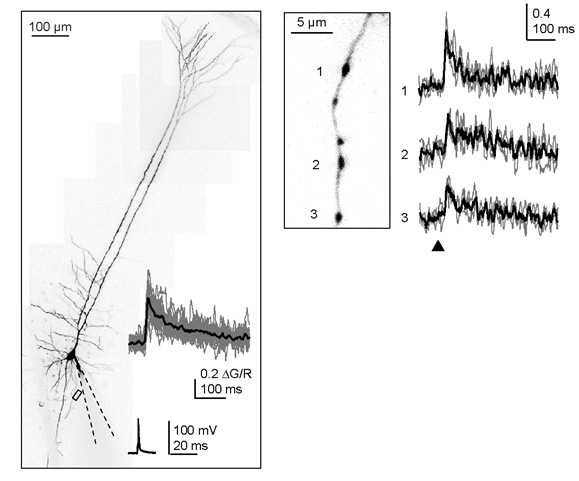
Figure 3. Presynaptic 2P- Ca2+ imaging at a layer 5 pyramidal neuron in an acute brain slice (left). Dashed lines indicate the patch-pipette. Ca2+ signals were induced by single action potentials and recorded from individual presynaptic terminals located on an axon collateral (box and right; modified from Bornschein et al., 2019)
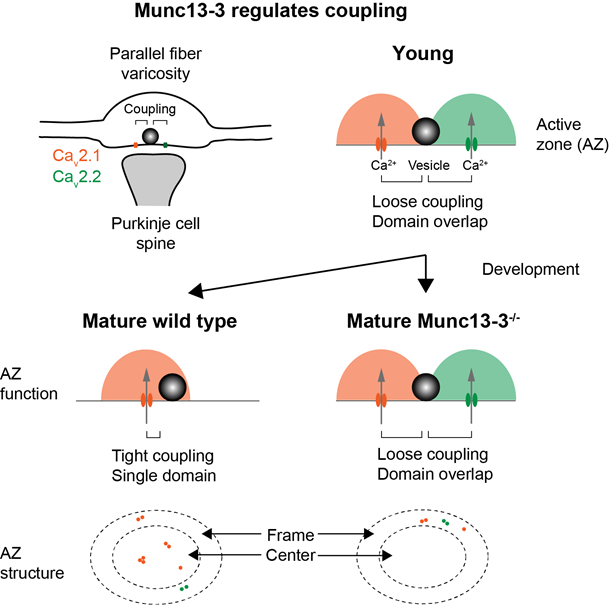
Figure 4. Numerical computer simulations were used to predict changes in active active zone topography accounting for functional maturation of the parallel-fiber to Purkinje cell synapse in wild-type and in the absence of the active zone protein Munc13-3 (from Kusch et al., 2018).
Ca2+ binding proteins: Mobility and Ca2+ binding kinetics
Ca2+ binds to Ca2+ sensing proteins that can be mobile or immobile. The diffusional mobility of proteins and binding to other proteins can be analyzed in synaptic structures by point-mode 2P- fluorescence recovery after photobleaching (FRAP; Fig. 5). Furthermore, by combining Ca2+ uncaging with presynaptic Ca2+ imaging it is possible to quantify the Ca2+ binding kinetics of Ca2+ sensor proteins (Fig. 6).
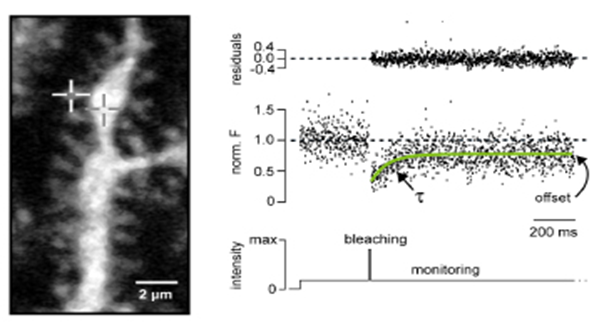
Figure 5. FRAP recordings. Dendrite and spines of a Purkinje neuron filled with dye labelled Calbindin (CB, left). To the right a FRAP recording from the spine marked by the white crosshair is shown. The offset indicates binding of CB in the spine. The time course of the recovery (τ) allowed quantifying the diffusion coefficient of CB (modified from Schmidt at al., 2005).
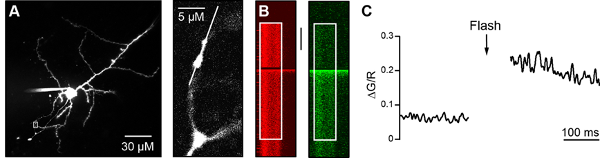
Figure 6. Ca2+ uncaging at neocortical boutons. A. Left: Two photon image of a pyramidal neuron equilibrated with a patch-pipette solution containing a Ca2+ indicator dye, a caged Ca2+ compound, and a red, Ca2+ insensitive dye. Right: Two presumed presynaptic terminals indicated by the box in the left panel. B. Line-scan from the boutons in A during a flash photolysis experiment. Red and green fluorescence were recorded. Scale bar, 100 ms. C. ∆G/R quantification of the area indicated by the white box in B. The flash artifact has been blanked.